
Structure, Architecture, and the Living Ecosystem Within Us
CONTENTS
• Introduction: The Hidden Majority
• Overall Shape and Arrangement
• The Cell Envelope
• The Cytoplasm and Its Contents
• External Appendages
• From Single Cells to Communities: Biofilms
• Bacterial Anatomy Across the Body
• Friend, Foe, and the Anatomy of Disease
• Conclusion: A Universe Within
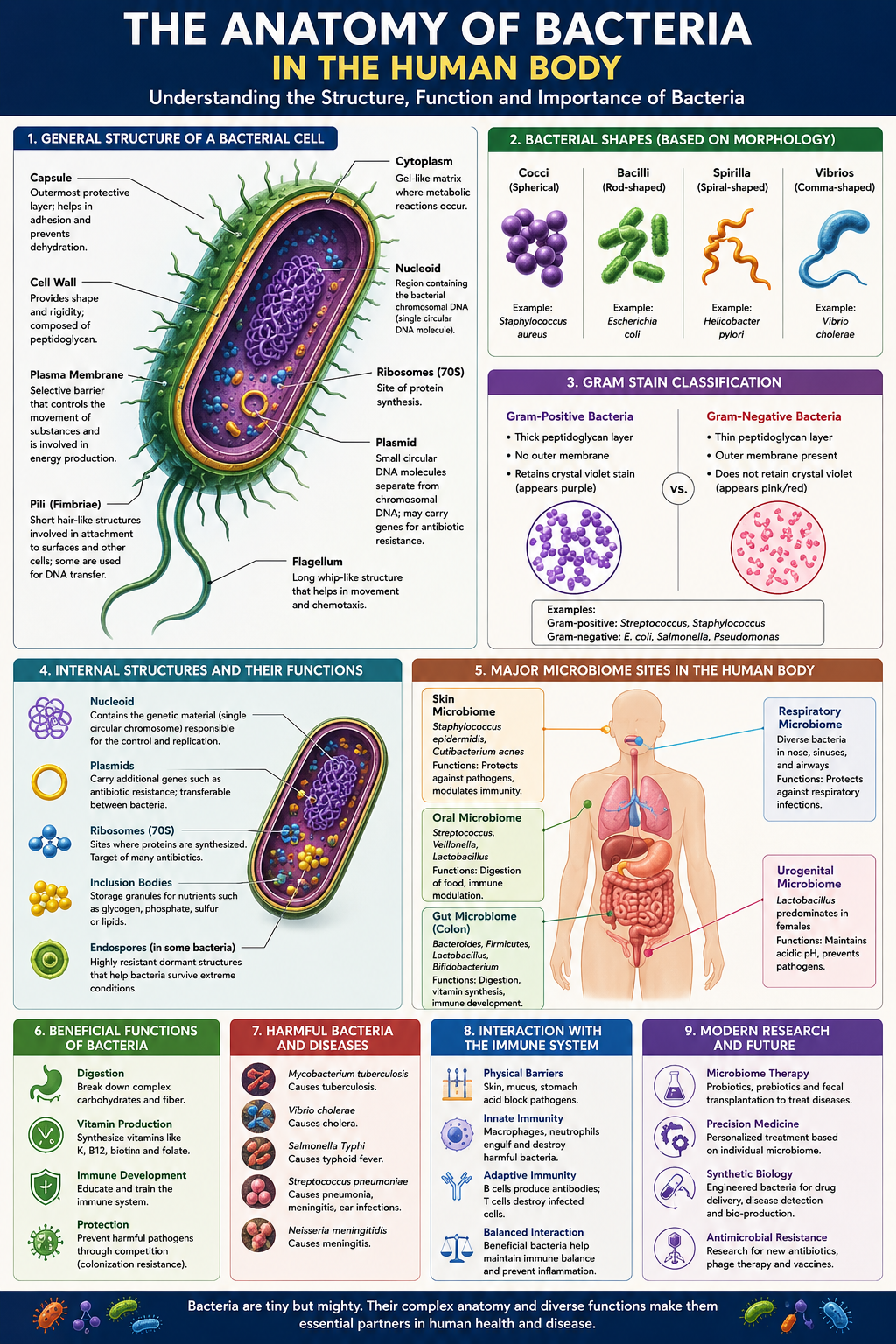
1INTRODUCTION: THE HIDDEN MAJORITY
If you could weigh every bacterium living in and on your body, the total would come to roughly one to two kilograms — about the weight of your brain. These organisms are not occasional visitors. They are permanent residents, numbering in the tens of trillions, and they outnumber or roughly match the human cells that make up your own tissues. You are, in a very real sense, a walking ecosystem.
This article examines what these bacteria actually are at the structural level. Before we can understand how gut bacteria help digest food, how skin bacteria defend against invaders, or how a harmful strain causes disease, we need to understand the anatomy of a single bacterial cell: its envelope, its interior, its appendages, and the architectural choices that make bacteria so successful. Anatomy, here, means the parts — what a bacterium is built from and how those parts fit together.
This tutorial walks from the outside of a bacterial cell inward: the layers of the envelope, the genetic material, the molecular machinery, the appendages used for movement and attachment, and finally how all of this plays out across the major bacterial communities of the human body — gut, skin, mouth, and beyond.
A crucial idea to carry throughout: bacteria are prokaryotes. This single word explains much of their anatomy. A prokaryote is a cell without a true, membrane-bound nucleus. The word comes from Greek roots meaning “before the kernel” — a reference to the missing nucleus, the “kernel” being the nut-like nuclear body found in our own cells. Human cells, by contrast, are eukaryotic (“true kernel”). This difference is the master key to bacterial structure.
1.1 Prokaryote versus Eukaryote at a Glance
The contrast between bacterial cells and human cells is the foundation for everything that follows. The table below summarises the major anatomical differences. Notice how much simpler the bacterial interior is — yet that simplicity hides remarkable sophistication.
Feature | Bacterial cell (prokaryote) | Human cell (eukaryote) | Nucleus | Absent; DNA floats in the nucleoid | Present; DNA enclosed in a membrane | Size | Typically 0.5–5 micrometres | Typically 10–100 micrometres | DNA shape | Usually one circular chromosome | Multiple linear chromosomes | Internal compartments | Few or none | Many (mitochondria, ER, Golgi) | Ribosomes | Smaller (70S) | Larger (80S) | Cell wall | Almost always present | Absent in animal cells | Reproduction | Binary fission | Mitosis / meiosis | A micrometre is one-thousandth of a millimetre. To picture the scale: if a typical human cell were the size of a basketball, a bacterium would be roughly the size of a grape sitting next to it. Hundreds of bacteria could line up across the width of a single human hair.
2OVERALL SHAPE AND ARRANGEMENT
Before dissecting the internal parts, it helps to recognise bacteria by their outward form. Shape (called morphology) is one of the first things a microbiologist notes under the microscope, and it is often a clue to identity. Bacterial shape is determined largely by the cell wall, which we will examine in detail in Section 3.
2.1 The Three Classic Shapes
Cocci (singular: coccus) are spherical or near-spherical. The bacteria that cause many skin infections and some forms of pneumonia are cocci. They are named like beads — round and compact.
Bacilli (singular: bacillus) are rod-shaped, like tiny capsules or sausages. Many gut residents, including the famous Escherichia coli, are bacilli.
Spirals are twisted or corkscrew-shaped. This group includes the curved vibrios, the rigid spirilla, and the flexible, coiled spirochetes — one of which causes syphilis.
Cocci do not always live alone. After dividing, they may stay attached in patterns that give further clues to identity: pairs (diplococci), chains (streptococci, like a string of beads), or irregular grape-like clusters (staphylococci). The prefix tells you the pattern; the root tells you the shape. Staphylococcus aureus, a common skin organism, is literally a “cluster-forming golden sphere.”
2.2 Why Shape Matters
Shape is not cosmetic. It affects how a bacterium moves through fluid, how efficiently it absorbs nutrients (a function of surface-area-to-volume ratio), how well it resists being swept away or engulfed by immune cells, and how it forms communities. A rod can swim directionally with more control than a sphere; a spiral can corkscrew through viscous mucus that would trap other shapes. Evolution has tuned bacterial geometry to specific niches in the body.
3THE CELL ENVELOPE: LAYERS OF THE BOUNDARY
The cell envelope is the collective name for the layered boundary separating the bacterial interior from the outside world. It is arguably the most important anatomical feature of a bacterium, because it does the work of containment, protection, sensing, and exchange all at once. Moving from inside to outside, the envelope can include the plasma membrane, the cell wall, and — in some bacteria — an outer membrane and a capsule.
3.1 The Plasma Membrane
The innermost layer of the envelope is the plasma membrane (also called the cell membrane or cytoplasmic membrane). It is a thin, flexible sheet built from a double layer of phospholipid molecules — the same basic design used by every living cell on Earth. Each phospholipid has a water-loving head and two water-fearing tails; the tails point inward toward each other, the heads face the watery environments inside and outside, and the result is a stable barrier just a few molecules thick.
In human cells, many specialised tasks are handed off to internal organelles. Bacteria have almost none of these, so the plasma membrane becomes a multi-purpose workbench. It is where energy is generated, where many molecules are built, and where the cell senses its surroundings. Embedded proteins act as pumps, channels, sensors, and enzymes.
Because a bacterium lacks mitochondria, the energy-harvesting reactions that a human cell runs inside its mitochondria are instead run on the bacterial plasma membrane itself. The membrane is studded with electron-transport proteins that pump charged particles to build up an electrical and chemical gradient — a stored form of energy the cell taps to make ATP, its molecular fuel currency.
3.2 The Cell Wall and Peptidoglycan
Just outside the plasma membrane sits the cell wall, a rigid mesh that gives the bacterium its shape and keeps it from bursting. Bacteria live in environments where water tends to rush inward by osmosis, swelling the cell. Without a wall, most bacteria would swell and pop. The wall acts like a corset — strong, slightly flexible, and load-bearing.
The signature material of the bacterial wall is peptidoglycan (also called murein). Its name describes its construction: “peptido” for short chains of amino acids, “glycan” for sugar chains. Long strands of sugar run in parallel, and short peptide bridges cross-link them together, producing a single enormous mesh-like molecule that wraps the entire cell like chain-mail. This material is unique to bacteria — human cells never make it — which is precisely why it is such a valuable drug target.
Penicillin and related antibiotics jam the enzymes that stitch peptidoglycan cross-links together. A growing bacterium keeps trying to build wall but can no longer seal it, so the weakened wall fails and the cell bursts. Because human cells have no peptidoglycan, the drug attacks the bacterium while leaving our own cells untouched — a textbook example of selective toxicity, where anatomy itself is the vulnerability.
3.3 Gram-Positive versus Gram-Negative: A Tale of Two Walls
In 1884, a Danish scientist named Hans Christian Gram discovered a staining method that splits nearly all bacteria into two great camps based on their wall architecture. The Gram stain remains one of the most important first tests in any microbiology lab, and the distinction it reveals is fundamentally anatomical.
Gram-positive bacteria have a thick peptidoglycan wall — many layers deep — sitting directly outside the plasma membrane. This dense wall traps a violet dye during staining, so these cells appear purple under the microscope. There is no outer membrane.
Gram-negative bacteria have only a thin peptidoglycan layer, but they add a second membrane outside it — the outer membrane. They lose the violet dye during washing and take up a pink counterstain instead. The outer membrane makes these bacteria harder for many antibiotics and immune molecules to penetrate.
Property | Gram-positive | Gram-negative | Peptidoglycan | Thick, many layers | Thin, few layers | Outer membrane | Absent | Present | Stain colour | Purple / violet | Pink / red | Endotoxin (LPS) | Absent | Present in outer membrane | Body examples | Staphylococcus, Streptococcus | E. coli, Salmonella | The space between the inner and outer membranes of a Gram-negative cell is called the periplasm or periplasmic space. Far from being empty, it is a busy compartment packed with enzymes that process nutrients, build the wall, and neutralise threats. Gram-positive bacteria, lacking an outer membrane, manage similar chemistry differently, often by secreting enzymes outward or anchoring them to the wall.
3.4 The Outer Membrane and LPS
The outer membrane of Gram-negative bacteria deserves special attention because of one molecule it carries: lipopolysaccharide, or LPS. As the name says, it is part lipid, part sugar. Anchored in the outer membrane and projecting outward, LPS forms a dense protective forest on the cell surface.
LPS is also called endotoxin, and it is medically infamous. When Gram-negative bacteria die and break apart in large numbers — for instance during a severe bloodstream infection — released LPS triggers the human immune system into a violent, system-wide alarm. The result can be the dangerous drop in blood pressure and runaway inflammation known as septic shock. Here anatomy and disease meet directly: a structural component of the bacterial envelope becomes a poison to the host.
3.5 The Capsule and Slime Layer
Some bacteria wrap themselves in an additional outermost coat made of a gel-like layer of sugars (and sometimes proteins). When this layer is thick, organised, and firmly attached, it is called a capsule. When it is looser and more diffuse, it is called a slime layer. Both are grouped under the term glycocalyx, meaning “sugar coat.”
A capsule serves two key roles in the body. First, it is camouflage and armour: its slippery surface helps the bacterium evade being recognised and engulfed by immune cells, making capsulated strains far more dangerous. Second, it is glue: the sticky coat helps bacteria adhere to surfaces — to teeth, to medical implants, to the lining of the gut — and to one another, which is the first step in building a biofilm (Section 6).
4THE CYTOPLASM AND ITS CONTENTS
Crossing the envelope, we arrive inside the cell, in the cytoplasm — the gel-like fluid that fills the bacterium. It is roughly 70 percent water, crowded with salts, sugars, proteins, and the machinery of life. Unlike the compartmentalised interior of a human cell, the bacterial cytoplasm is largely open-plan: a single shared space where most reactions happen side by side. Let us examine its main features.
4.1 The Nucleoid: DNA Without a Nucleus
The defining feature of a prokaryote is that its DNA is not locked inside a nucleus. Instead, the genetic material sits in an irregular region of the cytoplasm called the nucleoid (meaning “nucleus-like”). There is no membrane around it; the DNA is simply concentrated in this zone, in direct contact with the rest of the cell.
Most bacteria carry a single chromosome, and it is usually a closed loop — a circle of double-stranded DNA rather than the linear strands found in human cells. This single circular chromosome can carry several million base pairs of genetic code, all of it packed into a cell a few micrometres wide. To fit, the DNA is twisted and folded upon itself in a process called supercoiling, like coiling a long rope so tightly it occupies a fraction of its stretched-out length. Fully extended, the chromosome of a typical bacterium would be hundreds of times longer than the cell that holds it.
4.2 Plasmids: Optional Extra Genes
Besides the main chromosome, many bacteria carry small, separate loops of DNA called plasmids. A plasmid is like a portable accessory pack of genes — not essential for basic survival, but often extremely useful. Plasmids can be copied independently and, crucially, passed from one bacterium to another.
Plasmids are central to one of the great medical challenges of our era. Genes that make a bacterium resistant to antibiotics are frequently carried on plasmids. Because plasmids can be shared between cells — even between different species — resistance can spread through a bacterial population like a rumour through a crowd. A harmless gut bacterium can hand a resistance plasmid to a dangerous pathogen, arming it almost instantly. Understanding this small anatomical feature is essential to understanding why antibiotic resistance spreads so quickly.
4.3 Ribosomes: The Protein Factories
Scattered in their tens of thousands throughout the cytoplasm are ribosomes — tiny molecular machines that read genetic instructions and build proteins from amino acids. Every living cell has them, because proteins do nearly all the work of life: they form structures, drive chemical reactions, and carry signals.
Bacterial ribosomes are slightly smaller than human ribosomes. Scientists describe them by how fast they settle in a spinning tube: bacterial ribosomes are “70S” while human ones are “80S” (the S is a sedimentation unit, not a simple size measure). This difference is small but medically priceless. Several antibiotic families — including tetracyclines and erythromycin — bind specifically to the 70S bacterial ribosome and stall it, halting bacterial protein production while leaving the 80S human ribosome largely alone.
4.4 Inclusions and Storage Granules
Bacteria are thrifty. When nutrients are plentiful, many species stockpile reserves inside the cytoplasm as inclusion bodies — dense granules of stored carbon, phosphate, or other materials. When times grow lean, these stores are broken down for energy and building blocks. Think of them as the cell’s pantry: filled in good times, drawn down in bad. Some aquatic bacteria even store gas in tiny vesicles to adjust their buoyancy, floating up or sinking to find better conditions.
4.5 The Cytoskeleton
For decades bacteria were thought to be simple bags of fluid with no internal scaffolding. We now know this is wrong. Bacteria possess a cytoskeleton — a network of protein filaments that helps determine cell shape, guides the placement of the wall during growth, and pulls copied chromosomes apart during division. These filaments are the bacterial cousins of proteins that build the skeleton of our own cells, a reminder of the deep evolutionary kinship between all living things.
5EXTERNAL APPENDAGES: MOVEMENT AND ATTACHMENT
Many bacteria project structures outward from the envelope into the surrounding world. These appendages are tools for movement, attachment, and the exchange of genetic material. Three are especially important: flagella, pili, and fimbriae.
5.1 Flagella: The Propellers
A flagellum (plural: flagella) is a long, whip-like filament that a bacterium spins to propel itself through liquid. Remarkably, the bacterial flagellum is a true rotary motor — one of the very few known wheels in biology. Anchored in the envelope is a protein machine that rotates, turning the flagellum like a ship’s propeller. It can spin hundreds of times per second and reverse direction, driven by the same energy gradient stored across the plasma membrane.
Bacteria do not swim randomly. They sense chemicals in their environment — nutrients to approach, toxins to avoid — and steer accordingly, a behaviour called chemotaxis. By alternating smooth swimming with sudden tumbles that randomly re-aim the cell, a bacterium performs a biased random walk that carries it, on average, toward food and away from danger. The flagellum is the engine; the sensing system is the steering wheel.
The number and placement of flagella vary by species and are a clue to identity: some bacteria have a single flagellum at one end, some have a tuft, and some are covered all over like a hairy ball. In the human body, flagella let bacteria swim through fluids and mucus layers — helping beneficial microbes reach the gut lining and, unfortunately, helping pathogens penetrate protective barriers.
5.2 Pili and Fimbriae: Grappling Hooks and Bridges
Pili (singular: pilus) and fimbriae (singular: fimbria) are shorter, thinner, hair-like projections. The terms overlap in everyday use, but they group into two broad functions.
Fimbriae are numerous short bristles used mainly for attachment. They let a bacterium grip surfaces — the cells lining your bladder, your gut, your throat — so it can colonise rather than be washed away. For many pathogens, the ability to stick is the first and most essential step in causing infection.
Sex pili (or conjugation pili) are longer, fewer, and serve a different purpose: they connect two bacteria so DNA can pass from one to the other. This process, called conjugation, is one of the main routes by which plasmids — including those carrying antibiotic resistance — travel between cells. The pilus acts as a bridge and grappling line, drawing two cells together for the exchange.
5.3 Endospores: Survival Capsules
A handful of bacterial groups can perform an extraordinary feat when conditions turn hostile: they build an endospore. This is a dormant, armoured survival capsule formed inside the cell, containing a copy of the chromosome and a minimal kit of essentials, wrapped in tough protective coats. The rest of the cell then disintegrates, releasing the spore.
Endospores can survive boiling, drying, radiation, and many disinfectants for years — even, in some documented cases, for centuries. When favourable conditions return, the spore germinates back into an active cell. This is why hospital sterilisation must be so aggressive: ordinary cleaning will not destroy endospores. The bacteria that cause tetanus and botulism form endospores, which is part of what makes them so persistent and dangerous in the environment.
6FROM SINGLE CELLS TO COMMUNITIES: BIOFILMS
So far we have studied bacteria as individuals. But in the human body, bacteria rarely live alone. They cluster into communities called biofilms — organised mats of cells embedded in a self-produced slime. A biofilm is to a single bacterium what a city is to a person: a structured, cooperative, and far more resilient form of existence.
The dental plaque you brush off your teeth each morning is a biofilm. So is the film inside pipes, the slime on river stones, and the stubborn coating that forms on medical implants. Within a biofilm, bacteria stick to a surface and to each other using their capsules, fimbriae, and secreted polymers, building a matrix that shelters the whole community.
6.1 Why Biofilms Matter for Health
Biofilms are dramatically harder to eliminate than free-floating bacteria. The slime matrix slows the penetration of antibiotics and shields cells from immune attack. Bacteria deep inside a biofilm may also enter a slow-growing, dormant-like state in which antibiotics (which often target growth processes) work poorly. As a result, biofilm infections — on heart valves, catheters, artificial joints, and chronic wounds — are notoriously persistent and a major challenge in modern medicine.
Bacteria in a community coordinate their behaviour through chemical signalling called quorum sensing. Each cell releases small signal molecules; as the population grows, the signal concentration rises. Once it crosses a threshold — a “quorum” — the bacteria collectively switch on group behaviours such as building a biofilm or, in pathogens, launching an attack. It is a form of molecular voting that lets a population act only when its numbers are large enough to succeed.
7BACTERIAL ANATOMY ACROSS THE HUMAN BODY
Now we bring the anatomy of the single cell into the context of the whole body. The complete community of microbes living in and on us is called the human microbiome. Different regions of the body are distinct habitats — with different temperatures, acidities, oxygen levels, and food supplies — and each is home to characteristic bacterial residents whose anatomy suits the local conditions.
7.1 The Gut: The Great Fermentation Vat
The large intestine holds the densest bacterial community in the body — by far. It is a warm, low-oxygen, nutrient-rich environment, and the bacteria that thrive there are largely anaerobes: organisms that live without oxygen and would be harmed by it. Their plasma membranes run a different style of energy chemistry from the oxygen-using bacteria of the skin, an anatomical adaptation to a world without air.
These gut bacteria perform services we cannot do ourselves. They ferment dietary fibre that our own enzymes cannot break down, releasing short-chain fatty acids that nourish the cells lining the colon. They manufacture certain vitamins, including vitamin K and several B vitamins. They train and tune the immune system. And by occupying space and consuming resources, they crowd out invading pathogens — a protective effect called colonisation resistance.
7.2 The Skin: A Dry, Salty Frontier
The skin is a harsh habitat: comparatively dry, salty, acidic, and exposed. The bacteria that colonise it are adapted to these stresses, frequently relying on robust Gram-positive walls that resist drying out. They populate distinct micro-habitats — the oily zones beside the nose, the moist creases of the elbow, the dry expanse of the forearm — each a different anatomical neighbourhood.
Skin bacteria are not mere passengers. By occupying the surface and producing antimicrobial substances and acids, they make it harder for harmful organisms to gain a foothold. The healthy skin community is a living part of the body’s defensive barrier.
7.3 The Mouth: A Biofilm Showcase
The mouth is one of the most biofilm-rich environments in the body. Hundreds of bacterial species coat the teeth, tongue, and gums. On the hard, non-shedding surface of a tooth, bacteria build dental plaque — a classic, easily observed biofilm. When plaque bacteria ferment dietary sugars, they release acids that dissolve tooth enamel, which is the root cause of dental cavities. Here biofilm anatomy translates directly into a common human disease.
7.4 Other Niches
Every accessible surface of the body has its own community. The respiratory tract, the urinary and reproductive tracts, and the stomach each host bacteria adapted to local conditions — even the stomach, whose ferocious acidity was long thought sterile, harbours specialists able to survive there. In each niche, the anatomical features we have studied — wall type, capsule, appendages, oxygen chemistry — determine which bacteria can live where.
Body site | Conditions | Anatomical adaptation | Large intestine | Warm, low oxygen | Anaerobic energy chemistry | Skin | Dry, salty, acidic | Drying-resistant Gram-positive walls | Mouth / teeth | Wet, sugary, hard surfaces | Strong biofilm formation | Stomach | Highly acidic | Acid-survival mechanisms | Urinary tract | Flushed by fluid | Fimbriae for firm attachment |
8FRIEND, FOE, AND THE ANATOMY OF DISEASE
It would be a mistake to think of bacteria as enemies. The overwhelming majority of the bacteria in and on your body are harmless or actively beneficial. They are partners in digestion, immunity, and defence. A growing body of research links a balanced microbiome to health and a disturbed one to a range of conditions. We live in cooperation with our bacteria far more than in conflict.
Yet some bacteria do cause disease, and the anatomical features we have catalogued are precisely the tools they use. Drawing the threads together:
• Fimbriae and capsules let pathogens attach to and colonise body surfaces rather than being washed away.
• Capsules and certain wall features help them evade the immune system.
• Flagella help them penetrate protective mucus and reach vulnerable tissue.
• The LPS of Gram-negative outer membranes can trigger dangerous inflammation when bacteria die in large numbers.
• Plasmids and pili spread antibiotic resistance through bacterial populations.
• Biofilms shelter bacteria from antibiotics and immune attack, making infections persistent.
• Endospores let a few species survive sterilisation and lie dormant for years.
The same structures that make a beneficial gut bacterium good at sticking, surviving, and cooperating can, in a pathogen, become weapons. This is why understanding bacterial anatomy is not an academic exercise: it is the foundation of how we diagnose infections, design antibiotics, develop vaccines, and sterilise medical equipment. Every Gram stain, every penicillin tablet, every autoclave cycle is an application of the anatomy described in this article.
8.1 Anatomy as the Basis of Treatment
Notice a pattern running through this tutorial: nearly every successful antibacterial strategy targets a structure that bacteria have and humans do not. Penicillin attacks the peptidoglycan wall. Several antibiotics jam the 70S ribosome. The Gram stain reads the wall to guide treatment. Sterilisation must defeat the endospore. The bacterium’s anatomy is simultaneously the secret of its success and the map of its weaknesses.
9CONCLUSION: A UNIVERSE WITHIN
A single bacterium is, on close inspection, a marvel of compact engineering. Within a space a thousand times smaller than the dot on this letter ‘i’, it packs a complete envelope of protective layers, a coiled chromosome carrying millions of genetic instructions, tens of thousands of protein-building ribosomes, rotary motors for movement, grappling hooks for attachment, and the chemistry to power it all — all without a single internal compartment of the kind our own cells depend upon.
Multiply that single cell by tens of trillions, organise them into communities suited to every surface and cavity of the body, and you arrive at the human microbiome: a living, breathing, largely cooperative ecosystem that we carry with us from birth. To understand its anatomy is to understand ourselves a little more completely — not as isolated organisms, but as the densely populated, deeply interconnected ecosystems we truly are.
- Bacteria are prokaryotes — cells without a true nucleus — which explains most of their distinctive anatomy.
- The cell envelope (membrane, wall, and sometimes outer membrane and capsule) is the most important anatomical feature; the Gram stain reads its architecture.
- Peptidoglycan, the 70S ribosome, and the endospore are uniquely bacterial — and are therefore prime targets for antibiotics and sterilisation.
- Appendages (flagella, pili, fimbriae) drive movement, attachment, and gene transfer, including the spread of antibiotic resistance.
- In the body, bacteria live as biofilm communities tuned to each niche; most are beneficial partners, and the same structures that make them useful can, in pathogens, cause disease.
— End of Article —
Makoti Millennium Services Educational Series • June 2026



Be First to Comment