
How the Human Cell and Nervous System Generate, Route, and Interpret Electrical Signals
Executive Summary
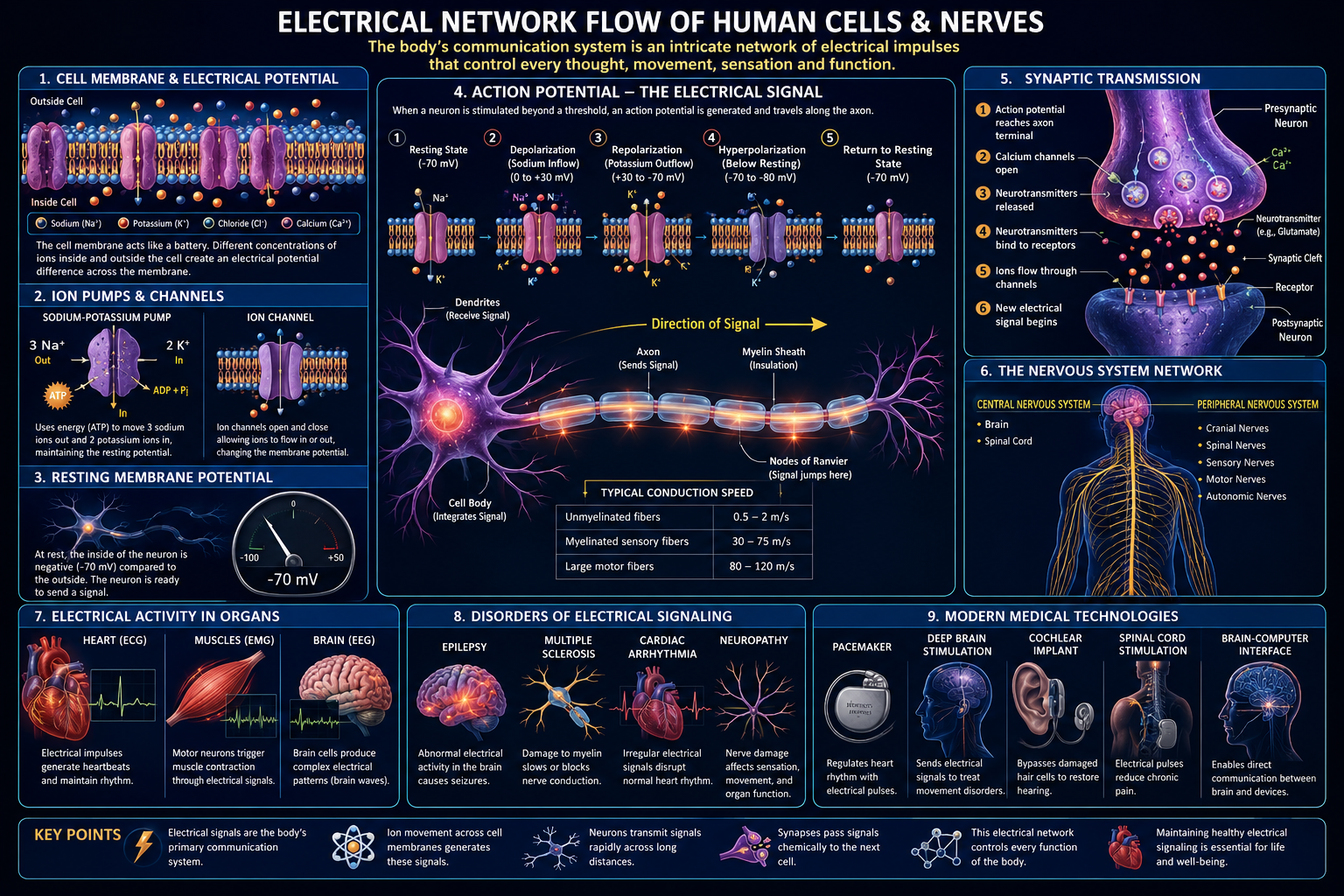
Every living human cell is, among other things, a small electrochemical battery. Across the thin, oily membrane that separates its interior from the outside world, the cell maintains a persistent voltage difference — typically around seventy millivolts — by pumping ions against their concentration gradients and by selectively opening and closing microscopic protein gates. Nowhere is this electrical machinery more elaborately exploited than in the nervous system, where roughly eighty-six billion neurons and their supporting glial cells form a densely wired network capable of carrying information at speeds exceeding one hundred metres per second.
This report treats the human body, deliberately, as an electrical network in the engineering sense: a system of conductors, capacitors, resistors, switches, and signal generators, obeying the same fundamental physics that governs any circuit — while being built from lipids, proteins, and ions rather than copper and silicon.
The report proceeds in ten sections, tracing the network from a single ion channel opening for a fraction of a millisecond up to the rhythmic electrical fields recorded from the surface of the scalp or chest.
Why an Engineering Frame Matters
The equations that predict a neuron’s resting voltage (Goldman-Hodgkin-Katz) and the decay of a signal along an axon (the cable equation) are the same mathematics used to design underwater telegraph cables in the nineteenth century — a rare case where physics, electrical engineering, and cell biology speak a genuinely shared language.
1. The Cell as an Electrical System
The plasma membrane is a phospholipid bilayer roughly five nanometres thick, an almost immeasurably thin film that nonetheless behaves, electrically, exactly like a parallel-plate capacitor. Its two faces — the ion-rich cytoplasm on one side and the ion-rich extracellular fluid on the other — act as the conductive plates, and the lipid interior acts as the dielectric insulator between them.
1.1 The Membrane as Capacitor
A biological membrane has a specific capacitance of approximately one microfarad per square centimetre, a value strikingly consistent across almost all animal cells, because it depends mainly on the physical thickness and dielectric properties of the lipid bilayer rather than on any specific protein content.
1.2 Ions as Charge Carriers
Unlike a copper wire, where free electrons carry current, the body’s electrical network is carried entirely by ions — sodium, potassium, calcium, and chloride — dissolved in water. Potassium is roughly thirty times more concentrated inside the cell than outside; sodium is roughly ten to twenty times more concentrated outside than inside. This asymmetry is actively created and continuously maintained.
1.3 Resistance and the Lipid Barrier
Pure lipid bilayer is an excellent insulator. Resistance drops dramatically the instant a channel protein opens a pore through the membrane, and it is the timed opening and closing of these pores that produces every electrical signal the body generates.
Order of Magnitude
A typical neuron’s resting potential is about -70 millivolts across a membrane only five nanometres thick — scaled to the thickness of a household wall, that gradient corresponds to roughly fourteen million volts per metre.
2. Ion Channels and Pumps: The Active Circuit Elements
If the membrane is the capacitor and insulator of the cellular circuit, ion channels and pumps are its transistors, switches, and batteries.
2.1 The Sodium-Potassium Pump
The sodium-potassium ATPase exports three sodium ions and imports two potassium ions for every ATP molecule consumed, working continuously against both gradients and contributing directly to the resting voltage since it moves unequal charge in each direction.
2.2 Voltage-Gated Channels
Each voltage-gated channel contains a voltage-sensing domain that physically moves when membrane voltage shifts, mechanically coupling to a gate that opens or closes — converting an electrical signal directly into a structural change.
2.3 Ligand-Gated and Leak Channels
Ligand-gated channels open in response to a chemical messenger rather than voltage. Leak channels are always open to some degree and set the baseline resting potential through a slow, continuous ion trickle.
2.4 Channelopathies
Mutations in ion channel genes underlie certain inherited epilepsies, cardiac arrhythmias such as long QT syndrome, and some forms of periodic paralysis.
3. The Resting Membrane Potential
A cell at electrical rest is not electrically inactive; it is actively holding a steady voltage, typically between -60 and -90 millivolts in neurons, through continuous, balanced pump and channel activity.
3.1 The Nernst Equation
The Nernst equation calculates the equilibrium potential for a single ion — the voltage at which electrical and chemical forces on that ion exactly balance. For potassium this predicts roughly -90 mV; for sodium, roughly +60 mV.
3.2 The Goldman-Hodgkin-Katz Equation
The Goldman equation extends this to multiple ions simultaneously, weighted by relative permeability — explaining why the resting potential of roughly -70 mV sits closer to potassium’s equilibrium value than sodium’s.
3.3 Why Rest Requires Constant Energy
The resting potential is a steady state maintained only through continuous ATP expenditure. Cut off energy supply, as in ischemia, and the resting potential collapses within minutes.
A Useful Analogy
Think of the resting potential as water held behind a dam by continuously running pumps, not water sitting still in a pond — the dam is leaky, and only tireless pumping keeps the level constant.
4. The Action Potential: A Travelling Electrical Pulse
The action potential is the nervous system’s fundamental unit of transmitted information: a brief, stereotyped, self-propagating reversal of membrane voltage that travels without losing amplitude.
4.1 Threshold and All-or-None
A stimulus must depolarise the membrane past roughly -55 mV to trigger a spike. Above threshold, the response is identical regardless of stimulus strength — a direct result of positive feedback between voltage and sodium channel opening.
4.2 The Five Phases
- Depolarisation: sodium channels open, voltage swings rapidly positive.
- Peak/overshoot: voltage briefly approaches sodium’s equilibrium potential.
- Repolarisation: sodium channels inactivate, potassium channels open and drive voltage down.
- Hyperpolarisation: potassium channels linger open, briefly overshooting rest.
- Refractory period: sodium channels stay inactivated, enforcing one-way travel.
4.3 The Refractory Period and Direction
The absolute refractory period guarantees one-directional propagation, since the just-fired region cannot fire again immediately.
4.4 Conduction Velocity
Speed depends on axon diameter and myelination — thicker axons offer less internal resistance, and myelin lets signals leap between exposed gaps rather than regenerating continuously.
5. Neurons as Circuit Elements
The physics of signal transmission along an axon is captured by the cable equation — the same mathematics developed for transatlantic telegraph cables in the 1850s.
5.1 The Axon as a Leaky Cable
An axon behaves like resistors and capacitors arranged along its length, with current leaking out through the membrane at every point. A passive signal would decay within a millimetre or two without active regeneration.
5.2 Myelin as Insulation
Myelin, produced by oligodendrocytes and Schwann cells, wraps axons in a multilayered spiral, increasing membrane resistance and decreasing capacitance in wrapped segments.
5.3 Saltatory Conduction
The signal jumps from node of Ranvier to node of Ranvier rather than regenerating continuously, increasing conduction velocity roughly tenfold over an unmyelinated fibre of the same diameter while conserving metabolic energy.
Clinical Relevance: Demyelinating Disease
Multiple sclerosis and Guillain-Barré syndrome both damage myelin, slowing or blocking saltatory conduction — a direct illustration of what happens when a biological cable’s insulation is stripped away.
6. Crossing the Gap: Synaptic Transmission
6.1 Electrical Synapses
Gap junctions connect adjacent cells’ cytoplasm directly, allowing near-instant current flow — ideal for extremely fast, synchronised firing such as cardiac muscle contraction.
6.2 Chemical Synapses
An arriving action potential triggers calcium influx, causing neurotransmitter-filled vesicles to fuse with the membrane and release their contents across a twenty-nanometre gap, converting the signal back to electrical form on the far side.
6.3 Excitation and Inhibition
A neurotransmitter’s effect can be excitatory or inhibitory depending on which channels it opens. A single neuron may integrate thousands of both inputs simultaneously.
6.4 Synaptic Plasticity
Synaptic strength can change with use — long-term potentiation and depression are the leading cellular candidates for how memories are physically stored, meaning learning itself remodels the brain’s electrical circuitry.
7. From Single Neurons to Networks
7.1 Circuit Motifs
Feedforward circuits pass signals in one direction; feedback circuits self-regulate and generate rhythm; lateral inhibition sharpens sensory contrast, as seen in retinal edge enhancement.
7.2 Oscillations and Synchrony
Neuron populations fire in coordinated rhythms — delta waves in deep sleep, alpha waves in relaxed wakefulness, gamma-band oscillations linked to binding perceptual features together.
7.3 The Brain’s Electrical Budget
Despite being about two percent of body mass, the brain consumes roughly twenty percent of resting metabolic energy, mostly restoring ion gradients after activity.
Network Scale
With around eighty-six billion neurons and hundreds of trillions of synapses, the brain’s connection count exceeds the transistor count of even the most advanced processors.
8. Reading the Network from Outside the Body
8.1 Electrocardiography (ECG/EKG)
The heart’s conduction system — sinoatrial node, atrioventricular node, bundle of His, Purkinje fibres — generates a stereotyped waveform whose P, QRS, and T features correspond to sequential atrial and ventricular activity.
8.2 Electroencephalography (EEG)
Scalp electrodes detect summed synaptic activity from large populations of aligned cortical neurons, used clinically for epilepsy diagnosis, sleep staging, and injury assessment.
8.3 Electromyography (EMG)
EMG records muscle fibre electrical activity, helping distinguish muscle disorders from the nerve disorders that supply them.
8.4 Nerve Conduction Studies
Stimulating a nerve at one point and recording downstream provides a direct measure of conduction velocity, useful for conditions like carpal tunnel syndrome.
9. When the Network Fails
9.1 Epilepsy
Excessive, hypersynchronised neuronal firing produces seizures, arising from channelopathies, structural lesions, or excitation-inhibition imbalance.
9.2 Cardiac Arrhythmias
Conduction disruptions, from AV node block to long QT syndrome, can range from benign to life-threatening ventricular fibrillation.
9.3 Peripheral Neuropathy
Diabetes and other conditions damage peripheral axons or myelin, typically starting in the longest nerves — a “glove and stocking” pattern.
9.4 Multiple Sclerosis
Immune attack on central myelin slows or blocks saltatory conduction, producing wide-ranging neurological symptoms.
9.5 Myasthenia Gravis
Autoimmune targeting of acetylcholine receptors weakens neuromuscular transmission, causing fatigable muscle weakness.
10. Frontiers: Bioelectricity Beyond the Nervous System
10.1 Bioelectric Signalling in Development
Embryonic cells maintain resting potentials and gap-junction communication before a nervous system exists; altering bioelectric patterns has been shown experimentally to redirect tissue fate.
10.2 Wound Healing and Regeneration
Injured tissue generates a measurable electric field that helps direct repair cell migration; regenerating species show distinct bioelectric signatures at injury sites.
10.3 Cancer and Bioelectricity
Some tumour cells display abnormal resting potentials, motivating early research into voltage-based normalisation approaches.
10.4 Bioelectronic Medicine
Vagus nerve stimulation, spinal cord stimulation, and deep brain stimulation directly modulate the body’s own electrical signalling as therapy, with closed-loop systems an active area of development.
Conclusion
From a single lipid molecule forming a five-nanometre insulating film, to eighty-six billion neurons whose synchronised rhythms are recordable from a scalp electrode, the human body runs on a continuous electrical logic — ion gradients as batteries, channels as switches, axons as insulated cables, synapses as relays, and coordinated neural firing as the oscillating fields read in an ECG or EEG.
As bioelectric principles extend into development, regeneration, and oncology, framing the body as an electrical network is proving not just a teaching device, but an increasingly literal and clinically actionable description of how living systems are organised — and how they might be repaired.
Further Reading and Reference Topics
- Hodgkin and Huxley’s model of the squid giant axon action potential
- Molecular structure and evolution of voltage-gated ion channel families
- Cable theory and its shared origins with nineteenth-century telegraphy
- Synaptic plasticity, long-term potentiation, and cellular models of memory
- EEG frequency bands and cognitive/sleep states
- Channelopathies in cardiology and neurology
- Developmental and regenerative bioelectricity
- Bioelectronic medicine and neuromodulation devices





Be First to Comment